
Designing Naturalistic Simulations for Evolving AGI Species
Christian Hahm
Department of Computer & Information Sciences, Temple University, Philadelphia, PA, U.S.A.
Keywords:
Adaptation, Artificial Evolution, Genetic Algorithm, Virtual Environment, Artificial Life, AI Embodiment,
Evolutionary Simulation.
Abstract:
This paper identifies basic principles for designing and creating evolutionary simulations in the context of
general-purpose AI (AGI). It is argued that evolutionary simulations which employ certain nature-inspired
principles can be used to evolve increasingly intelligent AGI species. AGI frameworks are particularly suited
for evolutionary experiments involving embodiment since they can operate arbitrary evolved bodies. Once a
designer manually defines a simulation’s initial conditions, each run is an automated exploration of a novel
subset of species. In this way, naturalistic simulations generate huge amounts of empirical data for evaluating
the robustness of AGI frameworks, along with many promising species that can be later instantiated in other
simulated environments or even physical robots for practical applications.
1 INTRODUCTION
Virtual simulation goes naturally with AI: since both
are implemented in digital computers, it is almost triv-
ial to interface the two. We should consider this a
huge benefit, since virtual environments (which, for
this discussion, can be referred to using other words
e.g., a “world”, “universe”, “reality”, “simulation”
etc.) are highly customizable by the designer. To per-
form “simulation” in the context of AI means:
1.) To design and create a virtual world (in a com-
puter, using a “simulator” software), and
2.) To embody one or more AI agents in that
world.
The purpose of simulation is to provide AI agents
a controlled environment in which to interact and
learn from their simulated experience, such that we
may study the agents (e.g., to observe their processes
under certain conditions) or utilize them in some real-
world application once they are sufficiently trained.
An agent is considered “embodied” in an envi-
ronment so long as it has sensorimotor mechanisms
(Wang, 2009), meaning sensors (i.e., tools on the
body which transduce environmental signals as inputs
to the mind) and actuators (i.e., tools on the body
controlled by output signals from the mind) in that
environment. The agent receives sensory signals to
monitor the environment, and can send output sig-
nals to move its body. Like a “brain-in-a-vat”, an AI
system treats sensory signals identically regardless of
whether they come from simulated virtual sensors or
real-world physical sensors.
Artificial general intelligence (AGI) is a term
which can be used to delineate cognitive-inspired
computer systems with full autonomy and general-
purpose ability. The possible applications of simu-
lation to AGI research are numerous, but mostly in-
volve optimizing the system on either the object level
or the meta level. In object-level testing, we embody
some agents and let them learn simulated tasks au-
tonomously. In meta-level testing, we vary config-
urable cognitive parameters to test various “personal-
ities” of AGI individuals on object-level tests. Cog-
nitive parameters may range from scalar control val-
ues (e.g., decision thresholds, attention decay, etc.) to
structures in the system’s architecture (e.g., buffers,
memory, etc.). The exact nature of cognitive parame-
ters depends on the selected framework.
Since AGI systems are general-purpose, they
should be capable of operating arbitrary bodies. In
considering embodiment, we can expand the notion
of parameters from cognitive values to include phys-
ical structures like the system’s morphology, sensors,
actuators, and physiology (bodily parameters). Any
parts of the system can be parameterized, except those
core aspects which we desire to remain static. When
aspects of the system’s mind and body are parame-
terized, it evokes the possibility of creating varied AI
“species”: agents which share a common core frame-
work of intelligence, but vary by their exact cognitive
296
Hahm, C.
Designing Naturalistic Simulations for Evolving AGI Species.
DOI: 10.5220/0011299600003274
In Proceedings of the 12th International Conference on Simulation and Modeling Methodologies, Technologies and Applications (SIMULTECH 2022), pages 296-303
ISBN: 978-989-758-578-4; ISSN: 2184-2841
Copyright
c
2022 by SCITEPRESS – Science and Technology Publications, Lda. All rights reserved

processes, bodily form, and sensorimotor capabilities.
Evolutionary simulations automate the process of
finding and optimizing such mind-body parameteri-
zations. Traditionally, evolutionary simulations prop-
agate species that maximize a quantitative “fitness”
function. Natural evolution admits no such fitness
function, and to simulate it requires a slightly dif-
ferent treatment than traditional genetic algorithms.
Instead of optimizing genomes to maximize a pre-
defined fitness function, “naturalistic simulations” re-
quire agents to prove their reproductive fitness in-
dependently, encouraging the evolution of intelligent
autonomy. If the selected AGI framework is flexible
and the simulated environment is sufficiently rich, the
resulting species should exhibit interesting adaptive
behaviors.
2 RELATED WORKS
Evolving autonomous machines is not a novel con-
cept; one of the earliest and most famous attempts at
elaborating this idea is mathematician John von Neu-
mann’s self-reproducing automaton. He hypothesized
a baseline level of complexity that allows the evolu-
tion of increasingly complex systems (von Neumann,
1966, p.78-80), realized by the three sub-processes
of reproduction and evolution: duplication, modifi-
cation, and instantiation of genomes. In his context,
the automaton system itself performs all three sub-
processes, including modifying the genome.
Though “AGI” is often construed as human-level
intelligence, our simulated agents will be more like
primitive animals than humans. They will not be ca-
pable of manually modifying their own genes; the
task of genomic modification belongs to the simula-
tion. As long as the agents handle reproduction au-
tonomously, the simulation design will handle the ac-
tual evolutionary processes behind the scenes. Game
engines such as Unity3D (Juliani et al., 2018) are the
ideal simulator software since they facilitate building
simulated physical worlds and arbitrary scripting.
(Holland, 1992) formalizes a methodology for ap-
proaching so-called “problems of adaptation”. This
work led to the popularization of genetic algorithms,
an evolutionary computing method by which to ex-
plore a space of evolving structures using concep-
tions of fitness, genetics, and reproduction. Genetic
algorithms are arguably the best approach for simu-
lating natural evolution, because they implement the
general principles that make natural genetics adap-
tive. Namely, genetic algorithms use “survival of the
fittest”, recombination, and random mutation to ex-
plore a certain space of evolving structures in search
of more optimal (i.e., “fit”) structures.
(Sims, 1994a; Sims, 1994b) use Holland’s genetic
algorithms to evolve various agent bodies and their
neural networks, including by facing the agents off
in direct physical competition (Sims, 1994a). The
agents are encoded using a highly compact and flexi-
ble genetic language in the form of directed network
graphs. Physical body parts contain sensors (e.g.,
contact sensor, light sensor) or actuators (e.g., rota-
tional force on joints), and each body part has a neural
circuit allowing for some level of distributed control.
(Soros and Stanley, 2014) propose four neces-
sary conditions for open-ended artificial evolution:
1&2.) new and existing individuals must be required
to meet some non-trivial minimum criterion (MC) be-
fore they can reproduce, so species do not degener-
ate into trivial-behavior automatons, 3.) individuals
must autonomously meet that MC, 4.) the complex-
ity of the individual’s genotype can grow unrestricted.
The naturalistic MC is the ability to reproduce, in-
cluding the requisite apparatus and motivation (Soros
and Stanley, 2014, p.2-3).
(Stranneg
˚
ard et al., 2020) establishes a “generic
animats” framework in which simulated organisms
are defined by homeostatic variables, sensors, pat-
terns, motor, actions, and reflexes. (Stranneg
˚
ard et al.,
2021) formalizes ecosystems containing inanimate
objects and organisms that have unique properties
(e.g., digital “animals” have energy levels, environ-
mental objects have certain chemical reactivity) and
common properties (e.g., all objects have mass).
For this discussion we will assume Wang’s work-
ing definition of intelligence: “the capacity of a
system to adapt to its environment with insufficient
knowledge and resources” (Wang, 1995; Wang, 2019,
p.13; p.17-18). According to this definition, the min-
imum requirement for intelligence is to operate un-
der an Assumption of Insufficient Knowledge and Re-
sources (AIKR). A system operating under AIKR: 1.)
uses constant computational resources, 2.) works in
real-time, and 3.) is open to new tasks regardless of
their content. In plainer terms, such a system is like a
living organism: it is a fast, finite agent that operates
effectively even under uncertainty, while constantly
learning from its experience so as to better achieve its
goals. Cognitive frameworks designed with AIKR in
mind may be flexible enough to permit the evolution
of complex adaptive behaviors.
Designing Naturalistic Simulations for Evolving AGI Species
297

3 SIMULATION DESIGN
3.1 Formalizing the Problem
Following (Holland, 1992, p.28,35), a given problem
of adaptation and the adaptive system to tackle it, in
our case evolving AI species in simulation, are de-
fined by the variables: α, Ω, T , ε, χ.
The variable α = {A
1
, A
2
, ...} is the set of all
genomes that can possibly be evolved; if the simu-
lation’s genetic language permits genomes to grow
without restriction, this set is infinite. An evolution-
ary run searches this set. Genome encoding should
include both cognitive parameters for the AGI system
and bodily parameters for its embodiment.
Ω = {ω
1
, ω
2
, ...} is the set of genetic operators
that can be used to modify genomes. A given op-
erator might perform some variation of crossover or
mutation.
T = {τ
1
, τ
2
, ...} is the set of possible reproduc-
tive plans, each of which is a sequence of operators
from Ω which can be used to incrementally traverse
α. One τ may be selected for the entire simulation
run, or the choice of τ could even be evolved.
ε = {E
1
, E
2
, ...} is the set of possible environ-
ments. Creating a concrete simulation requires speci-
fying the actual environment E ∈ ε.
Finally, χ represents a criterion to compare the
many possible plans in T .
So, to specify these variables is to determine the
possibilities of the evolutionary simulation. The con-
tents of plans in T depend on the operators avail-
able in Ω; for example, a plan could look like τ =
(ω
i
, ω
j
, ω
i
, ω
k
, ...). Operators in Ω depend on how
the designer specifies the genetic language for α.
Therefore, this discussion will mostly ignore the de-
pendent variables Ω and T , instead focusing on gen-
eral considerations for specifying α, E, and χ.
3.2 Embodiment
3.2.1 Atoms
One major design problem of virtual simulation is that
we have to manually specify its irreducible compo-
nents. There are no interacting molecules like there
are in reality, unless we program them. We want
to simulate a physical reality efficiently and with fi-
nite resources. We also want a flexible simulation,
which requires irreducible generic components that
can combine and interact in various ways.
The simulation can implement a notion of equal
exchange, where a finite number of virtual atoms un-
derlie all structures in the simulation. There could be
Figure 1: Atoms are the underlying virtual representation of
a simulation’s structures. In this mockup, the green block
is an environmental structure consisting of atoms. Agents
could consume the atoms to exchange them for homeostatic
maintenance, physical growth, or offspring.
various types of atoms with unique properties. The
atoms are abstract intermediary components, never
explicitly seen but used justify environmental struc-
tures in E and physical expression of body parts en-
coded by α (see Figure 1). Atoms could correspond
one-to-one with size-standardized polygons or voxels.
This prevents any structure from growing dispropor-
tionately compared to the others by enforcing equal
exchange.
While pure morphology can be grown incremen-
tally, polygon by polygon, some body parts must be
hardcoded with special-purpose functionality in the
form of modality-specific sensors, actuators, and mo-
tivation. For example, vision requires a special pho-
toreceptor body part which can render a partial view-
point of the scene and communicate it to the AGI sys-
tem. One way to handle this is to exchange atoms one-
to-one for pure structure (with some distinguishing
properties) whereas require a many-to-one exchange
for structures with special functionality. For a simple
example, two red and two green atoms could corre-
spond to a single photoreceptor, meaning a parental
agent will need to consume those atoms to instantiate
an offspring with a photoreceptor part. Agents would
return their body’s absorbed atoms to E upon death
(via simulated decomposition) for recycling.
The minimum E is an environment that hosts re-
producing (thus evolving) cognitive agents and re-
spects a conservation of materials. Once this is
achieved, E can be made more complex and inter-
active using frameworks like atoms, perhaps even
including genomes for other kingdoms of life (e.g.,
plants, fungus, etc.). Various “organisms” could reor-
ganize atoms in unique ways, in a simulated “circle of
life”. Non-cognitive entities like plants are not strictly
necessary, but without them the environment will be
simplistic and should not be expected to produce very
sophisticated or interesting agents.
SIMULTECH 2022 - 12th International Conference on Simulation and Modeling Methodologies, Technologies and Applications
298

3.2.2 Sensorimotor
In order to evolve sensorimotor, first the environment
E must simulate specific modalities (e.g., floating
odorants, sound waves), and the genomes in α should
be capable of expressing the corresponding modality-
specific sensors and actuators (e.g., olfactory organs,
visual organs, vocal cords, etc.). It is certainly worth
exploring ways to evolve arbitrary sensory modali-
ties. However, since sensors measure physical signals
(e.g., goodness of molecular fit, mechanical pressure,
wavelength of light, etc.) which do not exist by de-
fault in simulation, it seems necessary to explicitly
support certain modalities in the design of E and α.
Figure 2: Sensors such as tactile sensors on a 3D model
(left, middle) should be represented “topographically”,
where physically adjacent sensors are also represented adja-
cently, such as in a network (right). (Made with Blender, blender.org).
Natural sensorimotor neurons in the body bun-
dle then connect to the brain in spatially ordered
“topographic” mappings (see (Wolfe et al., 2006,
p.64,280,397)). This helps the agent understand the
spatial distribution of its sensations. Simulated sen-
sorimotor signals could be represented in a similar
way, where sensors of the same type record their sen-
sations in a topographic map. The map should be “to-
pographic” in that sensors which are physically close
and adjacent in the simulation are also represented ad-
jacently in the map (see Figure 2).
For example, it is quite intuitive to simulate simple
vision: render a full 2D visual image of the scene, and
treat each pixel of the image as a single visual sensor
(aka photoreceptor). The situation is muddier for the
other senses. More generally, topography can be de-
scribed using a network, where edges explicitly (as in
a graph, where nearby nodes are connected) or im-
plicitly (as in a convolutional neural network, where
neighboring nodes connect to a common node) rep-
resent adjacency. A network representation might be
useful for touch perception, which relies on unevenly
distributed sensors and may not be as amenable as vi-
sion to a 2D array representation. The AGI system
can then use the topography to selectively group and
process sensations (Wang et al., 2022).
3.3 Fitness and Reproduction
Traditionally with genetic algorithms there is an addi-
tional variable, µ, specifying the function by which to
measure an individual’s fitness. The simulation pro-
ceeds in discrete rounds, where after each round the
fitness of individuals is quantitatively measured us-
ing µ. Then, the genetic material of a few individuals
with the highest scores are combined to create many
offspring, whereas those with lower scores are culled.
In AI research, we would ideally select a µ that mea-
sures and thus optimizes for intelligence. However,
no widely-agreed µ exists to quantitatively measure
“intelligence”, amongst humans nor across species.
Besides, evaluating agent performance with a single
number is inflexible, since such evaluations are more
vulnerable to “hacky solutions” and much generality
is lost as species evolve towards optimizing only one
function.
In nature, “reproductive fitness” is a tautology.
There is no numeric fitness measure by which na-
ture decides reproduction, instead fitness is equiva-
lent to (or proven by) successful reproduction. Traits
which confer reproductive and survival advantages
in the current environment will tend to appear more
frequently in the populations than traits which pro-
vide relatively less advantage. Therefore, a naturalis-
tic simulation with an implicit µ should at the very
least yield increasingly prolific species. Reproduc-
tion, as in nature, can be done sexually or asexu-
ally. Asexual reproduction is simpler to simulate and
very quickly grows the population, though agents who
are selectively sexual might induce faster speciation
and evolved capabilities, as argued in (Canino-Koning
et al., 2017).
Although such a µ frees us from needing to explic-
itly measure fitness, it introduces a number of prob-
lems. First, all simulated agents need to meet the nat-
ural “minimum criteria” (Soros and Stanley, 2014):
they must be capable of reproducing autonomously.
“Capable” not only means physically able to perform
a reproductive action (as determined by the specific
simulation), but also requires the agents to be moti-
vated towards that action (i.e., to have a sex drive). In
biological contexts, bodily glands release hormones
that modulate reproductive motivation in the brain
(Wise, 1987; Fisher et al., 2006; Cummings and
Becker, 2012). A bodily origin of motivations in natu-
ral agents is interesting to notice in our context, since
it implies motivation evolves with the body.
There needs to be a tradeoff between consuming
atoms from the environment (e.g., food) and produc-
ing offspring. Consistent and balanced resource ex-
change is critical as it requires agents to earn their
Designing Naturalistic Simulations for Evolving AGI Species
299
success. The exact process of this tradeoff during re-
production depends on the simulation design and α.
One could simulate naturalistic processes like gesta-
tion (where a parent gets pregnant then gradually con-
sumes the relevant atoms to grow a baby), incuba-
tion, external egg fertilization, etc., though the bio-
logical details are not necessary to replicate, only an
energy/materials tradeoff to instantiate and grow the
offspring.
The choice of adaptive plan τ ∈ T for a given
simulation run is important, since it determines the
evolutionary trajectories through α. Yet, we do not
know the best τ to choose. χ is the quantitative cri-
terion for evaluating plans in T so as to find the best
τ. The χ is normally something like average fitness
of the population, ¯µ, so as to try various τ in appli-
cation and pick one which maximizes ¯µ, but since a
naturalistic µ is implicit, we cannot pick such a χ.
One decent option for χ is the total number of in-
dividuals produced by τ , which should result in sim-
ulations with many agents and hopefully more oppor-
tunities for beneficial evolution. However, any χ or τ
can be tried — more agents by time t does not nec-
essarily mean “more-intelligent” agents by time t. It
may be possible to treat χ implicitly like µ, leaving
the choice of τ to evolution by allowing the selection
of τ to vary depending on the species (in which case,
each A ∈ α should represent its selection of τ).
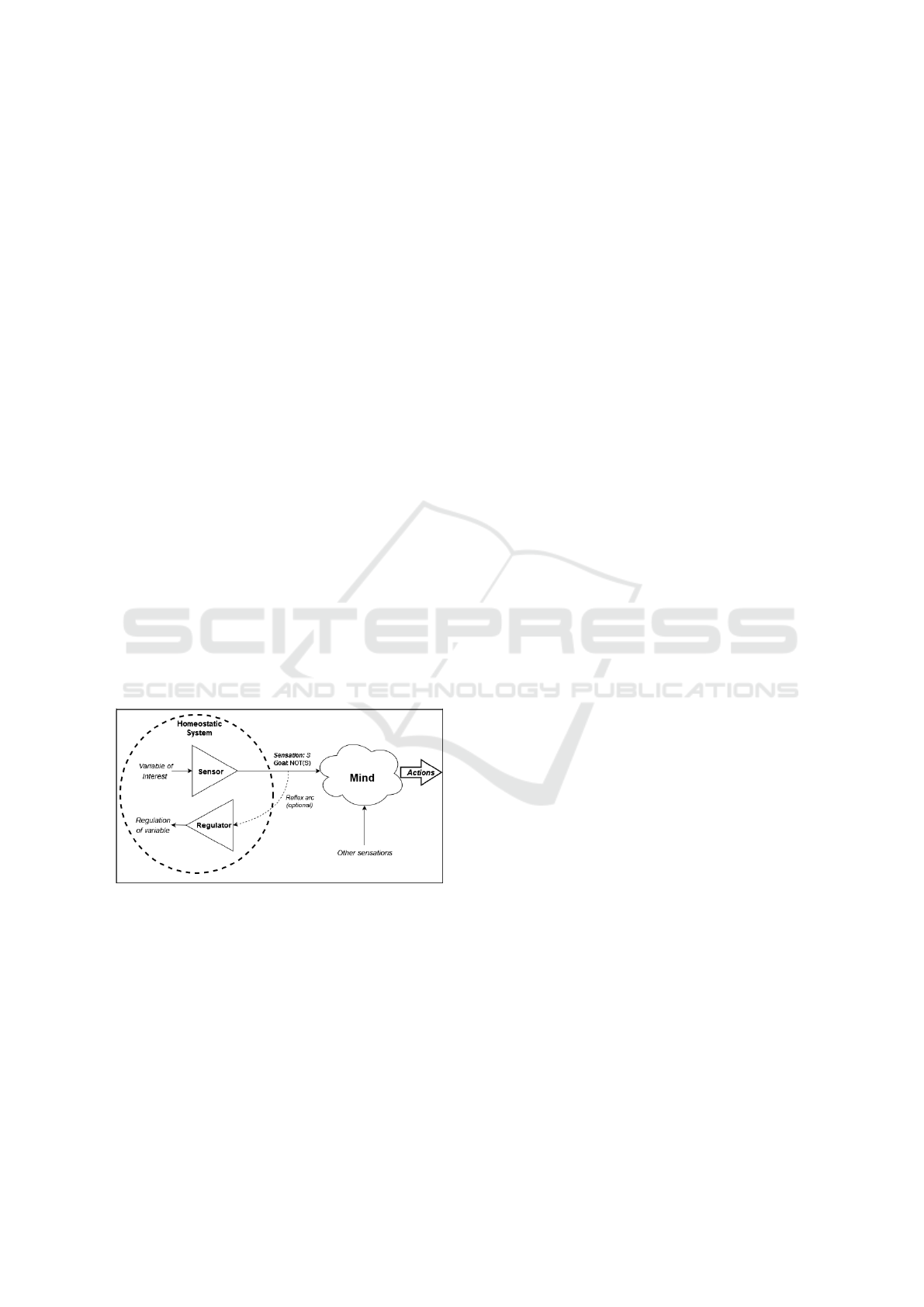
3.4 Homeostasis as Seed Motivation
Figure 3: Bodily systems entering homeostatic imbalance
may trigger sensations and motivation in the mind.
Homeostasis is the tendency of a system to maintain
bodily variables within an optimal range. The purpose
of homeostasis is to keep the system alive; whenever
there is a homeostatic imbalance, the body triggers
automatic reflexes and signals motivation to the mind
so as to fix the imbalance. (Tsitolovsky, 2015, p.3,4)
explains: “Needs are at the heart of motivations”,
and since homeostatic goals are endogenous (origi-
nating from within the agent’s own mind-body sys-
tem, rather than from an external user) they “turn [the
agent] into a subject with its own behavior”. Reflexes
only have very limited power to restore the body to
stability, depending on the severity and complexity
of the issue. This is why, in nature, both sensation
and motivation are signalled to the central nervous
system, so that the agent itself can restore homeosta-
sis by interacting with the environment (Craig, 2003;
Tsitolovsky, 2015).
Maintenance of the body as the agent’s source of
motivation makes perfect sense, as it keeps the agent
alive. Determining how agents source motivation is
extremely important since without motivation there is
no action. A small set of motivational signals from
the body is enough. Simple motivational “seeds” can
derive a wide range of additional motivations, causing
many complex behaviors to emerge depending on the
specific agent’s knowledge and capabilities (Hahm
et al., 2021). Evolving homeostatic systems on the
body can provide various seed motivation signals to
the agent. The more skilled the agent is at meeting
useful homeostatic survival needs, the more likely the
agent will survive to reproduce.
One could simulate homeostatic motivation
as in Figure 3, for example, within a body part
containing variable-specific sensors which activate
whenever some variable (e.g., heat, cold, pain)
veers too far from its acceptable range. The sensor
then signals both a sensation of the imbalance
S = Belief (sensation) and a negative desire to
alleviate it D = Goal(¬sensation). Alternatively,
a lack of sensation S = Belief (¬sensation) can
be paired with a positive desire to experience it
D = Goal(sensation). This model can account
for both avoidance behavior (e.g., polygons are
destroyed when they take enough damage, and
nociceptors detect damage. When the agent’s arm
mesh sustains a strong force, desire is signalled to
stop activation of the arm’s nociceptor):
{S, D} = {Belief(pain), Goal(¬pain)}
and approach behavior (e.g., green-atoms are the
agent’s store of energy, and have a property that is
detected by the mouth’s sweet-taste receptors when
eaten. When the agent’s green-atom stores drop
below critical levels, desire is signalled to activate the
mouth’s sweet-taste receptors):
{S, D} = {Belief(¬sweet), Goal(sweet)}
In the avoidance case, the agent is motivated to
stop an event (the pain, such as by withdrawing its
arm from the painful stimulus), whereas in the ap-
proach case, the agent is motivated to realize an
SIMULTECH 2022 - 12th International Conference on Simulation and Modeling Methodologies, Technologies and Applications
300

event (tasting sweet, such as by seeking and eating
green-atom). A sensation S alone is neutral informa-
tion, making the agent aware of the current situation.
However, by additionally signalling motivation D, a
homeostatic system influences the agent’s reaction to
S.
3.5 Evolving Intelligence (Implications
of AIKR)
We have conjectured that a naturalistic simulation
should produce increasingly prolific AGI species.
However, it is easy to see where this strategy will
fail. With a boxed-in environment and no energy con-
straints, agents will infinitely reproduce until the en-
vironment is overflowing with simple reproduction-
optimizers. In the context of AGI, we want to evolve
increasingly intelligent species, though prolific agents
help prevent global extinction. According to the as-
sumed working definition of intelligence, this means
we wish to improve each individual agent’s capacity
to adapt to its environment (i.e., its ability to learn
and execute skills on the object level) under AIKR,
using the evolutionary parameters we have available
(via genomic adaptation, on the meta level).
The condition of AIKR in the definition of intel-
ligence is an important nuance, because it moves us
slightly from the traditional viewpoint of intelligence
as an agent’s ability to adapt to an agent’s ability
to adapt under uncertainty and resource limitations.
So to evolve greater intelligence requires pressures in
the form of resource/energy limitations (i.e., insuffi-
cient resources) and uncertainty about E (i.e., insuf-
ficient knowledge). If the environment E is danger-
ously uncertain but the agent manages to survive it
and reproduce, then the resultant species might not be
reproduction-optimizers in the most simplistic sense,
but rather species who are reproduction-optimizers
despite the odds. Such species will necessarily ex-
hibit intelligent behaviors that are linked inextricably
to both their mind-body forms and the environment E
in which they evolved.
In other words, since we have no fitness func-
tion to pressure greater intelligence explicitly, agents
prove their superior capability in E independently by
surviving and achieving reproduction. If the environ-
ment is too gentle, there is no reason for species to
evolve better capabilities. If the environment is too
harsh, everyone goes extinct. One problem is, any ini-
tially “difficult” static pressures that we design in the
environment might be overcome by some specially-
adapted species, which would then explode in popu-
lation and plateau in complexity as the evolutionary
pressure is nullified.
This means the environment needs to exhibit
adaptive, even scaling, pressures on the global popu-
lation. In this regard, the agents are their own ideal
pressure, and they must be allowed to interact and
even (implicitly) forced to compete. As one species
improves their capability in E, another will be pres-
sured to keep up or risk extinction. Macro-scale
competition between species results in some species
adapting, then others, in a neverending back-and-
forth game of genetic improvements. The result is an
“arms races between and within species” (Dawkins
and Krebs, 1979), which is precisely what is needed
to prevent intelligence from plateauing.
The uncertainty arising from an uncountable
amount of complex evolving agent-environment and
agent-agent interactions in naturalistic simulation
would overwhelm traditional AI systems. For exam-
ple, reinforcement learning agents assume states are
repeatable, and so would fail in a naturalistic simula-
tion which never exactly repeats. On the other hand,
this treatment is possible with AGI systems working
under AIKR as they are fundamentally equipped to
deal with uncertainty.
3.6 Initial Conditions
Once the atoms and encoding for α are decided, the
actual initial state of environment E must be created.
This is up to creativity of the designer, with two no-
table constraints.
Firstly, E must be contained with finite (though
renewable) resources. Consider an environment that
is just an infinite 2D plane with agents and resources
on top if it. Not only would this provide agents with
infinite resources, but the agents might simply spread
out and never reproduce. Those that do would have no
reason to evolve greater abilities, as they could always
migrate to a fresh new location when they use up all
the simple resources in their current location. In con-
trast, organisms which are forced together will have
no choice but to interact with each other and compete
over the limited resources, thus pressuring impressive
evolution via the arms race phenomenon.
Secondly, E must be populated with one or more
initial organisms at t = 0. The organism must at min-
imum possess reproductive ability, motivation, and
embodiment, spending energy to do work. A starting
point is the most simplistic organism possible accord-
ing to your specification of α; perhaps a simple body
capable of movement, tactile sensors or a small eye,
asexual reproductive organ, and a simplified gastroin-
testinal system (mouth, stomach, etc.) to maintain en-
ergy levels. Such organisms should begin to fill up the
space of E, and the evolutionary takeoff begins.
Designing Naturalistic Simulations for Evolving AGI Species
301

4 SUMMARY
By selecting a general-purpose AI framework and ap-
plying nature-inspired principles in simulation, we
can explore a huge variety of AGI species and even
pressure them towards greater intelligence. Naturalis-
tic simulations provide insights into the selected AGI
model and could even yield interesting or impressive
species for real-world applications.
The first step is to define the atomic building
blocks of the simulation. These atoms provide justifi-
cation in the form of equal exchange for structures in
the environment E and the agent population. Atoms
should be fairly exchanged for energy expenditure
and the volume/function of body parts instantiated
from α, such as during offspring creation. The defi-
nition of “equal” in equal exchange might be arbitrary
and up to the designer, as long as the exchange rates
are coherent and invariant. Just as atom classes are
hardcoded, the simulated transduction of environmen-
tal signals to sensory signals seems to require hard-
coded functionality, depending on which modalities
the designer wishes to explicitly support.
The next step is to specify which aspects of the
AI system and body are parameterizable (and in turn,
evolvable). There are two overarching classes of pa-
rameters to evolve and encode in the genome: cog-
nitive parameters, which modify the selected AGI
model (e.g., its control system, architecture, decision
thresholds, etc.), and bodily parameters, which mod-
ify the agent’s embodiment (e.g., morphology, phys-
iology [including homeostatic systems], and sensori-
motor capabilities). As such, the selected AGI model
must expose certain cognitive parameters, whereas
the simulation designer specifies the available bodily
parameters.
It is important that parameters of both types co-
evolve since their interplay may be delicate when it
comes to the agent exhibiting coordinated behavior.
Parameterization is among the most important factors
to consider when designing an evolutionary simula-
tion, since it fundamentally constrains the range of
species that can possibly be evolved. The more gen-
eral and flexible these parameters are, the more op-
portunities for novel, interesting, and useful abilities
to evolve. On the other hand, too many options in
α (especially for parameters which drastically alter
the core functioning of the system) can impede the
search for high-performing species, as the simulation
may waste computational resources on offspring with
relatively incoherent mind-body parameterizations.
Motivation seeds are “externally” sourced by the
agent’s body parts during homeostatic imbalances,
though such homeostatic systems will likely only be
burdened by a species in exchange for increased sur-
vival and reproductive success. Sensation signals
keep the agent informed of its current homeostatic
outlook, while motivation signals sway the agent’s
behavior. At absolute minimum, reproductive moti-
vation should be guaranteed in every organism so as
to perpetuate evolution.
Despite hunger’s appearance as a motivation
unique to organic creatures, energy maintenance mo-
tivation (such as in the form of hunger) is likely also
essential in artificial agents, since working with finite
resources demands a work-energy tradeoff. There-
fore, an agent without hunger will quickly use all its
energy without replenishment and die before it can
reproduce. Energy limitations force an agent to be
smarter about how and when it acts so as to achieve
its goals.
In many cases it is implausible to numerically
measure performance on complex or abstract tasks.
This is especially true when it comes to measuring an
ill-defined concept like “intelligence”, which has been
interpreted in various ways. A naturalistic simulation
is open-ended, rejecting any single choice for fitness
function µ to evolve intelligent behavior. Instead, as
in nature, many capable species co-evolve when indi-
viduals of each species prove their own reproductive
worthiness.
Agents are pressured to evolve better adaptive ca-
pabilities when the environment contains dangerous
uncertainties (increasing the importance of operating
with insufficient knowledge) and their vital resources
are strained (increasing the importance of working
with insufficient resources). Relatively static environ-
mental resource pressures will become less relevant
as species evolve greater capabilities towards surviv-
ing in their environment. However, if the environment
is contained, the species themselves should exert scal-
ing adaptive pressures on each other as they evolve to
out-compete each other for the limited resources.
Though there are many open questions, natu-
ralistic simulation seems like a plausible way to
evolve AGI agents with various intelligent capabili-
ties. Species evolved in especially physically realistic
simulations could even be instantiated in real-world
robots designed analogously to the simulated bod-
ies. A simulation can be slowed down, allowing us
to observe agents interacting in real-time, or sped up,
to hasten the evolutionary process and explore new
species populations. Overall, naturalistic evolution-
ary simulations are a tool to automatically explore a
wide range of capable AGI species, collect empirical
data, and gain insights into both AI design and the
nature of “intelligence”.
SIMULTECH 2022 - 12th International Conference on Simulation and Modeling Methodologies, Technologies and Applications
302

REFERENCES
Canino-Koning, R., Keagy, J., and Ofria, C. (2017). Sex-
ual selection promotes ecological speciation in digi-
tal organisms. In ECAL 2017, the Fourteenth Euro-
pean Conference on Artificial Life, pages 84–90. MIT
Press.
Craig, A. D. (2003). A new view of pain as a homeostatic
emotion. Trends in neurosciences, 26(6):303–307.
Cummings, J. A. and Becker, J. B. (2012). Quantitative as-
sessment of female sexual motivation in the rat: Hor-
monal control of motivation. Journal of neuroscience
methods, 204(2):227–233.
Dawkins, R. and Krebs, J. R. (1979). Arms races be-
tween and within species. Proceedings of the Royal
Society of London. Series B. Biological Sciences,
205(1161):489–511.
Fisher, H. E., Aron, A., and Brown, L. L. (2006). Roman-
tic love: a mammalian brain system for mate choice.
Philosophical Transactions of the Royal Society B: Bi-
ological Sciences, 361(1476):2173–2186.
Hahm, C., Xu, B., and Wang, P. (2021). Goal generation
and management in nars. In International Confer-
ence on Artificial General Intelligence, pages 96–105.
Springer.
Holland, J. H. (1992). Adaptation in natural and artificial
systems: an introductory analysis with applications to
biology, control, and artificial intelligence. The Uni-
versity of Michigan Press.
Juliani, A., Berges, V.-P., Teng, E., Cohen, A., Harper, J.,
Elion, C., Goy, C., Gao, Y., Henry, H., Mattar, M.,
et al. (2018). Unity: A general platform for intelligent
agents. arXiv preprint arXiv:1809.02627.
Sims, K. (1994a). Evolving 3d morphology and behavior
by competition. Artificial life, 1(4):353–372.
Sims, K. (1994b). Evolving virtual creatures. In Pro-
ceedings of the 21st annual conference on Computer
graphics and interactive techniques, pages 15–22.
Soros, L. and Stanley, K. (2014). Identifying necessary con-
ditions for open-ended evolution through the artificial
life world of chromaria. In ALIFE 14: The Fourteenth
International Conference on the Synthesis and Simu-
lation of Living Systems, pages 793–800. MIT Press.
Stranneg
˚
ard, C., Engsner, N., Ferrari, P., Glimmerfors,
H., S
¨
odergren, M. H., Karlsson, T., Kleve, B., and
Skoglund, V. (2021). The ecosystem path to agi. In
International Conference on Artificial General Intelli-
gence, pages 269–278. Springer.
Stranneg
˚
ard, C., Xu, W., Engsner, N., and Endler, J. A.
(2020). Combining evolution and learning in com-
putational ecosystems. Journal of Artificial General
Intelligence, 11(1):1–37.
Tsitolovsky, L. E. (2015). Endogenous generation of goals
and homeostasis. In Anticipation: Learning from the
past, pages 175–191. Springer.
von Neumann, J. (1966). Theory of Self-Reproducing Au-
tomata. University of Illinois Press. Edited and Com-
pleted by Arthur W. Burks.
Wang, P. (1995). Non-Axiomatic Reasoning System: Ex-
ploring the Essence of Intelligence. PhD thesis, Indi-
ana University.
Wang, P. (2009). Embodiment: Does a laptop have a body?
In Goertzel, B., Hitzler, P., and Hutter, M., editors,
Proceedings of the Second Conference on Artificial
General Intelligence, pages 174–179.
Wang, P. (2019). On defining artificial intelligence. Journal
of Artificial General Intelligence, 10(2):1–37.
Wang, P., Hahm, C., and Hammer, P. (2022). A model of
unified perception and cognition. Frontiers in Artifi-
cial Intelligence, 5.
Wise, R. A. (1987). Sensorimotor modulation and the vari-
able action pattern (vap): Toward a noncircular defini-
tion of drive and motivation. Psychobiology, 15(1):7–
20.
Wolfe, J. M., Kluender, K. R., Levi, D. M., Bartoshuk,
L. M., Herz, R. S., Klatzky, R. L., Lederman, S. J.,
and Merfeld, D. (2006). Sensation & perception. Sin-
auer Sunderland, MA.
Designing Naturalistic Simulations for Evolving AGI Species
303
